Interests • Functional Basis of Demographic Trade-offs • Seed Dispersal & Seedling Regeneration • Plant-Soil Feedbacks & Soil Microbiome • Biodiveristy & Change in Nebraska's Forests
Research: Functional Basis of Demographic Trade-offs
Trade-offs and the Distributions of Tree Species
Individual organisms face trade-offs in terms of how energy and resources are allocated during a lifetime. For example, a juvenile tree growing in the understory of a closed-canopy forest allocates resources to produce leaves: Those leaves may be inexpensive to produce and therefore require frequent replacement, but have great photosynthetic capacity enabling fast growth in height or, alternatively, those leaves may be stronger and more resistant to damage, and thus longer-lived, but more costly to produce.
Trade-offs are also important for understanding the distributions of tree species across heterogeneous environments. If we think in a hierarchical way about the ecological processes determining tree species distributions, we know that species distributions are an outcome of demographic processes (growth, survival, and recruitment). Demographic processes, in turn, are an outcome of the interaction between a species ecological “strategy” and the abiotic and biotic environment. These strategies represent functional (physiological & morphological) solutions to evolutionary optimization problems involving resource conservation, acquisition, and allocation. There are trade-offs among these strategies.
My laboratory has been using this hierarchical framework to investigate how trade-offs affect the distribution of tree species in a hyper-diverse rain forest of Borneo. One explanation for the exceptionally high tree species richness in these forests is that species coexist by occupying slightly different ecological niches and hence trade-off performance in different habitats. But how are plant species sorted among habitats that they could conceivably occupy in the absence of interspecific competition?
We work in a large forest dynamics plot in Borneo (Lambir Hills National Park in Malaysian Borneo) with collaborators from Smithsonian ForestGEO, Harvard University, and Malaysia's Forest Research Corporation. (See this fascinating history of ForestGEO, demonstrating the importance of global interdisciplinary collaboration.) We have been examining possible demographic and functional mechanisms that may generate the striking soil-correlated tree species distributions observed at Lambir (Russo et al. 2005, Baillie et al. 2006, Russo et al. 2007).

Our research has shown that these soil-correlated distributions can be explained in part by performance-based ecological sorting of species among soils. Sorting may be partly mediated by an interspecific demographic trade-off between a species' ability to tolerate low-resource conditions by having conservative demographic responses (slow growth and low mortality) vs. its ability to respond quickly to increases in resource availability (fast growth). Mortality risk for species with intrinsically fast growth is greater on the poorest soil, which is consistent with the trade-off hypothesis (Russo et al. 2008).
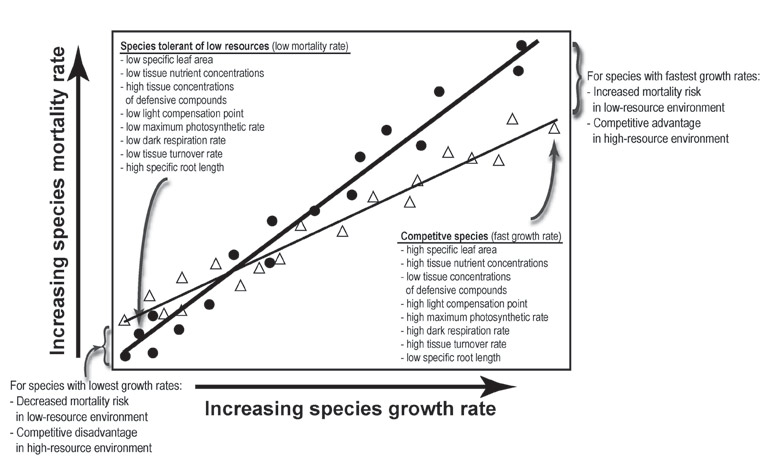
This interspecific demographic trade-off can be seen as an emergent property resulting from functional constraints related to physiology, morphology, and resource allocation expressed at the individual level. We are developing models relating this demographic trade-off to the possible functional mechanisms underlying it (Russo et al. 2010), by measuring in situ variation in ecophysiological and resource-allocation traits of tree species and relating this variation to plant performance, thereby linking plant function to its consequences for the structure and diversity of forest communities. However, these linkages are complex, since functional trait variation does not always correlate with demography (Paine et al. 2015).
However, we recently used tree-level demographic data to derive two parameters, tolerance and responsiveness, that describe the variation in species' resource allocation strategies thought to underlie the growth-mortality trade-off. We use these parameters to examine the determinants of the interspecific growth-mortality trade-off in 11 tropical forests. The trade-off was observed in all of the forests with mild disturbance regimes, which also had a more even representation of resource allocation strategies (tolerance and responsiveness). On the other hand, the trade-off was not observed in many disturbance-prone forests, which did not contain a diversity of resource allocation strategies. Thus, while the growth-mortality trade-off is common, it is not a general framework for understanding the community structure of tropical forests (Russo et al. 2020). Also see this blog for a "behind the scenes" look our paper.
In a review paper (below) led by former PhD student Bailey McNichol, we generalized these functional trade-offs and their implications for plant species' capacity for geographic and habitat range shifts. Together with former postdoctoral fellow, Sam Worthy, we are following up on this work, integrating microclimate variation, to understand large-scale patterns in geographic and habitat ranges of North American tree species.
- McNichol, B. and S.E. Russo. 2023. Plant species’ capacity for range shifts at the habitat and geographic scales: A trade-off-based framework. Plants 12(6):1248, doi.org/10.3390/plants12061248
Trade-offs in carbon allocation to growth, storage, and defense
Trees allocate endogenous resources, such as carbon and nitrogen, to different functions. Thinking hierarchically, the growth and survival of a tree is likely to be strongly influenced by how it allocates these resources, such as using carbon to make new leaves for light capture versus to make new roots for nutrient uptake. We have been focusing on the trade-offs in the allocation of carbohydrates to growth, storage, or defense, as a key mechanism underlying tree species distributions along soil fertility gradient. Trees store carbohydrates (e.g., in parenchyma cells in stems and roots), which may enable greater tolerance of environmental stresses. Using a seedling reciprocal transplant experiment with a defoliation treatment using 13 dipterocarp tree species, we tested the hypothesis that variation in storage carbohydrates is a unifying currency by which stresses due to nutrient- and water-limitation and due to tissue loss from herbivory and physical damage are translated into survival differences.

We found that the amount of stored carbohydrates was a strong correlate of both growth and survival of seedlings, patterns that have rarely been shown experimentally. Storage was negatively associated with leaf growth rate, suggesting an allocation-based trade-off. Survival of defoliation increased strongly with greater levels of stored carbohydrates. However, tree species with contrasting soil specializations did not differ significantly in their average allocation to storage.
This study is one of the first to experimentally demonstrate that allocation to storage carbohydrates in trees exhibits a trade-off relationship with growth and is a unifying currency by which environmental stressors are translated into the survival differences that lead to differences in species’ distributions along soil gradients.
With collaborators in a NIMBioS working group, we are using a dynamic energy budget modeling approach to develop a whole-tree physiological process model to predict tree growth and survival in response to environmental variation. See Couvreur et al. 2018 and Ledder et al. 2020 for two publications resulting from our math-biology modeling working group.
Trade-offs in leaf structure and longevity

Once carbon has been allocated to a particular function, a tree has many options for how to build that tissue or organ. The structure of leaves, for instance, plays an important role in their carbon-fixation function, with thin, short-lived leaves generally having faster rates of photosynthesis than thicker, more robust, longer-lived leaves. Unlike leaves in temperate deciduous forests, which are shed during the winter, the lifespans of tree leaves in the tropical forests can be very long and vary dramatically among tree species.
Leaf lifespan was one of the few leaf functional traits that correlated significantly with species growth and mortality rates in Bornean saplings, suggesting that it the duration of return on investments in leaves may be an important component of trade-offs related to resource allocation. The Russo lab is investigating these ideas by quantifying leaf functional trait variation in combination with long-term monitoring of leaf lifespans in Bornean forest (Russo & Kitajima 2016, Russo et al. 2010). In our most recent census, we found trees of several species that still retained leaves that were marked in our original census in 2004, making those leaves over 11 years old!